Dos vestígios faunísticos provenientes da intervenção arqueológica realizada no Mercado Velho de Palmela
Frederico Tátá Regala
Introdução
O Dr. António Rafael Carvalho teve a amabilidade de facultar ao signatário deste trabalho a possibilidade de analisar o significativo conjunto de vestígios faunísticos coligidos durante as escavações arqueológicas que promoveu no Mercado Velho de Palmela.
Segundo os dados fornecidos por este arqueólogo, os materiais em foco provêm de um depósito resultante da acumulação de lixos produzidos em ambiente urbano, cronologicamente enquadráveis nos séculos XIV e XV. Trata-se portanto de uma lixeira onde se conservaram testemunhos da actividade humana em Palmela, constituindo evidentemente uma importante fonte de informação. O estudo da componente faunística irá contribuir para uma leitura mais completa sobre alguns aspectos do quotidiano das populações que povoaram a região, particularmente no tocante à alimentação e aos modos de exploração dos recursos animais.
O trabalho realizado sobre os referidos materiais encontra-se na sua fase inicial, atendendo a que não foi ainda concluído o tratamento preliminar do espólio, nomeadamente as tarefas de lavagem, contagem geral e triagem. Ainda assim, foi possível proceder paralelamente à identificação taxonómica de algumas das peças, em particular as mais completas ou com melhor preservação dos aspectos de diagnose. Como resultado do trabalho até aqui realizado, obteve-se já uma visão abrangente sobre o elenco das espécies animais representadas.
Para além das referências quanto à metodologia adoptada e às fontes de informação mais regularmente utilizadas, é prematura a discussão de grande parte das informações obtidas. Apenas se avançam algumas cogitações ligadas à identificação taxionómica de três peças específicas de carnívoros, duas das quais particularmente problemáticas do ponto de vista da sistemática.
O espólio – aspectos gerais
O espólio foi recebido por lavar, em sucessivos lotes de sacos que foram posteriormente acondicionados em contentores. A totalidade dos materiais ocupa actualmente 10 contentores com 26x50x32 cm, que se encontram provisoriamente armazenados no depósito da Cripta Arqueológica do Castelo de Alcácer do Sal.
As peças que foram entretanto lavadas apresentam-se geralmente bem conservadas, mantendo por vezes as texturas superficiais praticamente inalteradas, exceptuando a habitual patine cromática resultante do contacto com os sedimentos. O grau de fragmentação é muito diverso, variando de pequenas esquírolas até peças ósseas inteiras. Em muitos casos evidenciam-se marcas de corte e outras que serão analisadas de modo detalhado posteriormente.
Critérios metodológicos
Para a identificação das peças osteológicas recorreu-se à bibliografia habitualmente utilizada pelos investigadores nesta matéria, sendo de realçar as seguintes obras: Schmid, E. (1972); Amorosi, T. (1989); Izquierdo, E. (1986); Cornwall, I. (1956); Cohen, A & Sergeantson, D. (1996); Boessneck, J. (1969); Pales & Lambert (1971); Pales & Garcia (1981). Foi adicionalmente utilizada a osteoteca pessoal do signatário, que corresponde a uma colecção constituída fundamentalmente por materiais resultantes da recolha e processamento de animais atropelados junto às estradas e linhas ferroviárias, lançados por agricultores, caçadores e pecuários para o fundo de algares, carcaças encontradas em praias, etc. Os exemplares de peixes foram, em regra, adquiridos no mercado. Deste modo, foi reunida uma significativa amostragem de exemplares da fauna doméstica e selvagem comum. Foi criado um ficheiro associado à colecção que reúne os dados descritivos das condições da recolha, modo de preparação, critérios de diagnose taxionómica e informações quanto ao animal recolhido, incluindo a respectiva fotografia.
Os exemplares malacológicos foram identificados com base nos catálogos da especialidade e em algumas publicação específicas, sendo de salientar: Macedo, M. et al. (1999) e Nobre, A. (1931/40). Também neste caso foi utilizada a colecção privada do signatário. Ocasionalmente, foram obtidas informações específicas junto do Dr. João Regala, licenciado em Biologia Marinha e Pescas pela Universidade do Algarve, a quem cumpre agradecer.
As medições realizadas sobre algumas das peças foram obtidas com craveira de precisão (escala de vernier), seguindo os métodos e critérios preconizados por Driesch, A. (1976) e por Desse J., Chaix, L. & Desse-Berset, N. (1986). Em alguns casos concretos foram obtidas medidas adicionais em conformidade com as descritas pelos diversos autores consultados.
A orientação metodológica genérica das tarefas foi influenciada pelas obras de alguns autores, entre os quais: Davis, S. (1989); O’Connor, T. (2000); Reitz, E. & Wing, E. (1999); Chaix, L. & Méniel, P. (2001).
A terminologia taxionómica usada no presente trabalho segue as adoptadas por alguns dos autores consultados, nomeadamente: Macedo et al. (1999) ao nível da malacologia (que segue Sabelli et al., 1992); a de Cardoso (1993a) para os mamíferos, a de Cohen & Sergeantson (1996) para as aves, e a de Izquierdo (1988) para os peixes.
Os materiais arqueológicos não tinham sido alvo de qualquer tratamento após a respectiva exumação. Careciam portanto de lavagem, tarefa essa de que se tem encarregue o signatário pessoalmente. Até ao momento, foi lavado cerca de um terço do total do espólio recebido, utilizando a água canalizada e com o auxílio de uma escova dentária. As peças são depois dispostas em tabuleiros plásticos revestidos com papel de jornal e deixadas a secar em ambiente interior, fora do alcance da luz solar directa. Deste modo, a secagem processa-se lentamente reduzindo as possibilidades de deformação ou fissuração em peças mais friáveis.
Após a secagem, faz-se uma triagem prévia com a separação em lotes distintos segundo a natureza dos materiais. De cada saco obtêm-se subconjuntos mediante a abjunção da componente malacológica e de peças mais frágeis, passíveis de se danificar pelo contacto com outras mais resistentes e pesadas. Procede-se à replicação exacta das etiquetas de modo a manter todas as embalagens individualmente referenciadas.
Os elementos mais completos e aqueles que mantêm bem preservadas as características consideradas importantes para a identificação sistemática das respectivas espécies são, em alguns casos, embalados individualmente para que se possa passar de imediato a uma primeira abordagem do foro taxionómico. Deste modo, rapidamente se obtiveram peças facilmente identificáveis, recorrendo à bibliografia disponível e aos materiais da osteoteca seguramente classificados. Depois de identificadas, essas peças passam, por sua vez, a servir de modelos comparativos para fragmentos de interpretação menos evidente. Daí a opção por esta sequência de trabalho, que permite ir juntando um complemento comparativo importante, o qual vai aumentando gradualmente, em simultâneo com a fase de lavagem. Este conjunto será por fim utilizado para a revisão detalhada dos restantes materiais.
No caso de ossos do esqueleto apendicular, são isolados os que tenham pelo menos uma epífise completa ou apenas ligeiramente danificada. Não se está a tentar ainda a identificação da maior parte das vértebras nem de costelas. Alguns dentes e elementos mandibulares são isolados para amostragem. As conchas mais facilmente reconhecíveis como os mexilhões, berbigões e amêijoas são contadas e embaladas logo após a secagem.
Também as peças aparentemente invulgares ou que exibam qualquer vestígio de afeiçoamento que sugira a sua eventual utilização como artefactos, são desde logo separadas e etiquetadas. Assim, tem sido possível trabalhar paralelamente a um nível mais especializado com alguns dos testemunhos. Encontram-se nesta situação certas peças de identificação mais delicada, para as quais é exigível uma pesquisa rigorosa.
Em casos pontuais procedeu-se já à obtenção de medidas para comparação com os dados biométricos disponíveis na bibliografia. Esta tarefa só será porém realizada de modo sistemático numa fase mais avançada do trabalho.
Listagem das espécies identificadas até ao momento
MAMÍFEROS
Artiodáctilos
Suídeos
Sus domesticus / scrofa L., 1758 – porco/javali
Cervídeos
Cervus elaphus L., 1758 - veado
Bovídeos
Bos taurus L., 1758 - boi/vaca
Capra hircus L., 1758 - cabra
Ovis aries L., 1758 - ovelha
Perissodáctilos
Equídeos
Equus caballus L., 1758 – cavalo
Equus sp.
Carnívoros
Canídeos
Canis cf. lupus L., 1758 – lobo ?
Canis familiaris L., 1758 - cão
Felídeos
Felis cf. sylvestris Schreber, 1777 – gato bravo ?
Felis catus L., 1758 – gato doméstico
Lynx pardina (Temminck, 1824) – lince ibérico
Lagomorfos
Leporídeos
Oryctolagus cuniculus (L., 1758) - coelho
AVES
Faisanídeos
Gallus gallus L., 1758 – galo / galinha
PEIXES
Gadiformes
Merluccius merluccius (L., 1758) – pescada
Perciformes
· Sparídeo indeterminado
MOLUSCOS
Gastrópodes
Docoglossa
Patellidae
Patella sp. - lapa
Vetigastropoda
Trochidae
Monodonta lineata (da Costa, 1778)
Neotaenioglossa
Littorinidae
Littorina littorea (L., 1758) - borrelho
Neogastropoda
Muricidae
Hexaplex trunculus (L., 1758) - búzio
Bivalves
Mytiloida
Mytilidae
Mytilus edulis L., 1758 - mexilhão
Pterioida
Pectinidae
Pecten maximus (L., 1758) – vieira
Chlamys varia L., 1758
Ostreoida
Ostreidae
Ostrea edulis L., 1758 – ostra redonda
Crassostrea angulata (Lamarck, 1819) – ostra portuguesa
Veneroida
Cardiidae
Acanthocardia sp.
Cerastoderma edule (L., 1758) - berbigão
Solenidae
Solen marginatus Pulteney, 1799 - lingueirão
Scrobiculariidae
Scrobicularia plana (da Costa, 1778) - lamejinha
Veneridae
Venus verrucosa L., 1758 – pé de burro
Tapes decussatus (L., 1758) – amêijoa cristã
CRUSTÁCEOS
Decápodes
Majidae
Maja squinado Herbst, 1788 - santola
Identificações consideradas problemáticas: os casos de lobo/cão e de gato-bravo/doméstico:
As questões concretas referentes à determinação específica das peças do esqueleto de lobo/cão e de gato-bravo/doméstico mostram-se de facto pertinentes, atendendo a que estas espécies são, na realidade, respectivamente as mesmas do ponto de vista biológico. Não existe, nem num caso nem no outro, incompatibilidade reprodutora uma vez que dos cruzamentos entre lobos e cães ou entre gatos-bravos e domésticos resulta descendência fértil. Existe simplesmente uma divergência evolutiva relativamente recente, resultante da domesticação. Por outro lado, a identificação de uma ou a outra variante fenotípica tem, sem dúvida, diferentes repercussões na respectiva interpretação em contexto histórico-arqueológico. Isso justifica a responsabilidade que existe em procurar uma determinação fiável. Assim, pretende-se expor os critérios de diagnose considerados nestes dois casos.
- O possível lobo - Trata-se de uma única peça dentária, nomeadamente um canino superior direito com marca de corte na extremidade da raiz, que provocou a ablação do apex. Provém do quadrado U 5, U.E. 14a , conotável com o século XV segundo A. R. Carvalho. Foi comparado directamente com alguns exemplares homólogos de cães domésticos actuais, sendo um destes de grande porte, embora de raça indeterminada. As dimensões globais do exemplar em causa revelaram-se significativamente superiores a este último. Os diâmetros tirados na base da coroa dentária: mésio-distal (14,7mm) e vestíbulo-lingual (8,7mm) indicam que a referida peça dentária provém de um grande canídeo. Será de um cão doméstico ou do seu provável agriótipo selvagem, o lobo europeu?
Está demonstrado que as dimensões dentárias dos cães domésticos são proporcionalmente inferiores às dos lobos. A domesticação reflecte-se, entre outros aspectos, na redução do tamanho dos dentes, especialmente os caninos e os carniceiros (Clutton-Brock, J., 1969:306). É bastante expressivo o gráfico apresentado por esta investigadora (fig.1), especialmente considerando que inclui exemplares de Great-Dane e Bloodhound. Segundo Brewer, D. (2001:15), em raças de cães domésticos corpulentos que ultrapassaram a compleição dos lobos, as dimensões dentárias mantêm-se proporcionalmente inferiores, mesmo considerando as linhagens antigas. Este autor cita inclusivamente o caso do São-Bernardo e do Irish-Wolfhound, cujos dentes permanecem relativamente pequenos em comparação com o tamanho atingido pelo crânio. Posto isto, saliento que as dimensões da peça em foco se enquadram próximas dos limites máximos obtidos em Canis lupus actuais adultos da Colecção do Museu Bocage (Pires et al., 2001/2002), e também de jazidas quaternárias portuguesas (Cardoso, 1993a).

Atendendo aos considerandos referidos sobre a proporcionalidade entre as dimensões dentárias e a compleição física do respectivo animal, facilmente se conclui que um dente correspondente a um grande lobo, na variante doméstica teria que corresponder a um cão de compleição extremamente avantajada. De notar também que a maioria das raças muito corpulentas de cães actualmente conhecidas foram geradas em tempos mais recentes do que os contextos de onde proveio esta peça. Até mesmo no âmbito das raças grandes antigas, é necessário que se leve em consideração que os animais agigantados que hoje podemos admirar são normalmente fruto de transformações sofridas com base em selecção artificial moderna e principalmente contemporânea. Este fenómeno deve-se sobretudo à actividade de criadores que produzem exemplares para exibição em concursos e outros eventos públicos.
Quanto às grandes raças portuguesas tradicionais, conforme se pode ver pelo gráfico da fig. 2, apenas o Rafeiro Alentejano ultrapassa ligeiramente (+2 cm) a estatura do Lobo Ibérico, ao nível das espáduas. Os pesos máximos referidos na bibliografia citada são de 50 kg em ambos.

O lobo é tradicionalmente um animal acossado pelo homem. A má reputação deste animal e os prejuízos que causa ao pastoreio levam a que seja perseguido e morto, situação que invariavelmente ocorreria na região de Palmela, onde existiam extensas áreas florestais, com coutadas reais. A presença, entre os materiais em estudo, de outras peças pertencentes a animais selvagens (veado e lince), são reveladores da actividade venatória.
Os lobos capturados poderiam ser levados para a povoação como troféus de caça, para aproveitamento da pele, e também devido às características medicinais ou de natureza supersticiosa que eram atribuídas a alguns dos seus órgãos. Cito, a título de curiosidade, um interessante exemplo obtido a partir de uma obra da primeira metade do século XVIII, do Doutor Francisco da Fonseca Henriques, médico de D. João V, que refere o seguinte: “Hum cinto de pelle de lobo hé remedio da gotta coral.”; “A carne de lobo prezerva das bruxas os meninos que a comerem.”; “O fígado de lobo tem virtude occulta para curar a gotta coral.” As frases transcritas encontram-se em: “ÍNDICE ALFABETICO – Das coufas mays notaveys defte livro.”
Considerando todas as informações recolhidas, acrescidas das evidências biométricas obtidas até ao momento, referentes a cães domésticos e a lobos, não resta alternativa senão considerar provável a hipótese de se estar na presença de uma peça pertencente a um lobo de compleição avultada. Ressalva-se porém que será importante reunir mais alguns elementos biométricos em caninos superiores de cães domésticos de grande corpulência, para que a identificação possa efectivamente ser dada como inequívoca.

2 – Canino superior direito de um cão doméstico actual cujo crânio tem 212 mm de comprimento côndilo-basal e 121 mm de largura bizigomática.
- O possível gato-bravo - De entre os materiais analisados, o gato-bravo poderá estar igualmente representado por uma única peça proveniente de: Q, U6-14a. Trata-se de um úmero esquerdo incompleto, fracturado na extremidade proximal, mas com a epífise distal inteira. Os critérios considerados para a classificação desta peça são semelhantes aos relatados para o caso anterior. Na realidade, o osso em questão foi comparado directamente com um homólogo de gato doméstico e com um exemplar de Felis sylvestris cuja identificação foi atestada através da análise biométrica da dentição. A diferença de dimensões é razoavelmente clara. O diâmetro máximo da epífise distal (21,3mm) encontra-se acima da média obtida por Cardoso (1993a), com base em sete exemplares de Felis sylvestris actuais. À semelhança do que sucede com os cães, os gatos domésticos são em regra mais pequenos e menos robustos que os seus parentes selvagens (Barret, P. & Macdonald, D., 1993; Castells, A. & Mayo, M.1993). A existência actual de gatos domésticos de grandes dimensões deve-se, na maior parte dos casos, a indivíduos anómalos ou a modificações rácicas recentemente induzidas. Acresce o facto de existir uma peça homóloga e outras biometricamente atribuíveis a gato doméstico, provenientes do mesmo contexto arqueológico, o que demonstra a coexistência dos dois morfotipos.
Considerando que a pele de gato-bravo foi utilizada e teve valor comercial, este animal era alvo de caça. A Lei de Almotaçaria de D. Afonso III de 26 de Dezembro de 1253 (vide Antunes, M. T., 1989), estabeleceu os preços das peles de varias espécies selvagens, incluindo a de gato-bravo. Embora referente a período histórico mais recuado, tal diploma legal constitui um testemunho da exploração deste animal.
O conjunto dos dados analisados autoriza a considerar a possibilidade de a peça em causa pertencer a Felis sylvestris, espécie que habitou a Serra da Arrábida até muito recentemente. Não é de excluir, também, a possibilidade de uma “hibridação”, ou melhor dizendo, miscigenação, situação comum que, neste caso, parece ser corroborada pelo aspecto um pouco grácil da diáfise (7,2 mm). Deve porém tomar-se em linha de conta que, de acordo com Smith (1969), a fusão da epífise distal do úmero se dá, nos gatos, até aos sete meses de idade e que a fusão da epífise proximal é bastante mais tardia (um a dois anos). Ora, no exemplar de Palmela, é ainda possível distinguir a linha de sutura do epicôndilo medial, na face posterior, junto ao foramen supracondylicum, o que significa que este osso não teria ainda completado a fase de crescimento. Tal aspecto é consubstanciado pela porosidade na parede óssea da região proximal da diáfise, que lhe atribui uma feição juvenil.
Apesar do exposto, é de admitir que a reduzida amostragem de valores referentes a gatos domésticos afecta a possibilidade de serem firmemente estabelecidas as conclusões pretendidas, especialmente considerando que a separação entre gatos domésticos e gatos selvagens está melhor demonstrada ao nível do crânio e em material dentário (Kratochvil & Kratochvil, 1979). Em conclusão, de acordo com os dados considerados, pode afirmar-se que a peça em causa confere com as homólogas de gato-bravo (Felis cf. sylvestris), sem que possa excluir-se, em absoluto, a hipótese de se tratar de um gato-doméstico muito corpulento ou de um animal resultante do cruzamento entre estes.

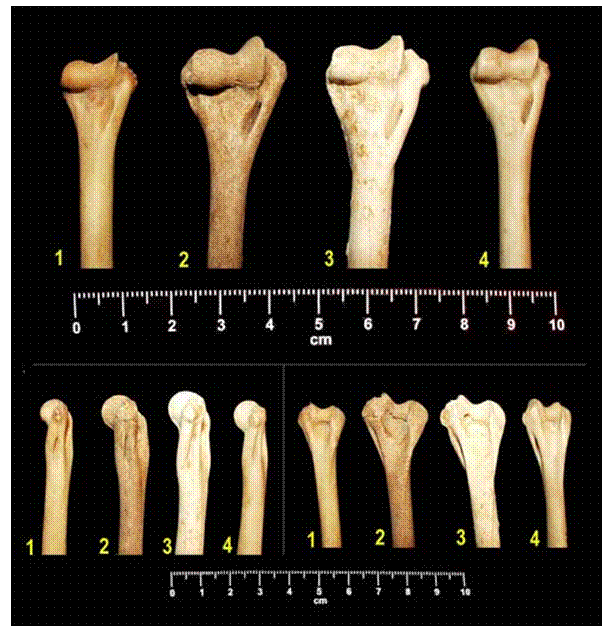
Fig. 4: Úmeros esquerdos de gatos: 1 – F. catus (Quadrado U6, U.E. 9); 2 - Felis cf. sylvestris (Quadrado U6, U.E. 14a); 3 - F. sylvestris (Algar da Manga Larga – Planalto de Santo António); 4 - Felis catus (Alcácer do Sal – exemplar actual adulto).
A mandíbula de lince
De entre os materiais osteológicos provenientes do quadrado S2-3, foi identificada a hemimandíbula esquerda de um lince. Foi recolhida na unidade estratigráfica 30, que corresponde a um nível de lixeira cronologicamente integrável entre os finais do século XIV e inícios do XVI (Carvalho, 2005). Trata-se de uma peça que apresenta fracturas antigas cuja coloração não difere da observável na globalidade das superfícies. O ramo mandibular ascendente está quebrado, assim como as duas extremidades do côndilo articular e a do processo angular. Os dentes incisivos estão ausentes mas a restante dentição permanece nos respectivos alvéolos. O dente canino encontra-se quebrado, faltando-lhe boa parte da coroa dentária. O terceiro pré-molar está completo e sem desgaste evidente. O quarto pré-molar apresenta indícios de fractura ante-mortem, nomeadamente um lascamento invasor do esmalte dentário na face vestibular, que alcançou o ramo dianteiro da raiz e que apresenta polimento da superfície de fractura causado pelo posterior uso do dente. O protocónido apresenta elevado grau de desgaste que poderá estar também relacionado com uma fractura no extremo coronal, ocorrida em vida, cujas arestas teriam sido ulteriormente boleadas durante a mastigação. O metacónido mostra também algum desgaste da superfície oclusal. O primeiro molar sofreu fractura extensiva, com arestas vivas, que resultou na perda da maior parte da coroa, restando apenas uma parte da face vestibular e a raiz. Em todos os dentes se verifica fissuração profunda.
A dentição plenamente desenvolvida revela um animal adulto, o que facilita as tarefas de comparação biométrica com outros exemplares. Para este efeito foram utilizados os dados obtidos por Garcia-Perea (1991), nomeadamente os relativos às populações actuais de lince-ibérico e de lince-boreal das regiões Centro, Norte e Este da Europa. As medições realizadas na peça em apreço seguiram os critérios descritos pela referida autora:
LC1 (=C1-DMD) – diâmetro mésio-distal do canino inferior ao nível do alvéolo (entre o bordo alveolar anterior e o posterior do C1);
AC1 (=C1-DVL) – diâmetro vestíbulo-lingual do canino inferior ao nível do alvéolo (entre o bordo alveolar vestibular e o lingual do C1);
LP3 (=P3–DMD) – diâmetro mésio-distal do terceiro pré-molar inferior, entre os pontos mais oral e mais aboral do cíngulo;
AP3 (=P3–DVL) – diâmetro vestíbulo-lingual do terceiro pré-molar inferior, entre os pontos labial e lingual mais laterais do cíngulo;
LP4 (=P4-DMD) – diâmetro mésio-distal do quarto pré-molar inferior, entre os pontos mais oral e mais aboral do cíngulo;
AP4 (=P4-DVL) - diâmetro vestíbulo-lingual do quarto pré-molar inferior, entre os pontos labial e lingual mais laterais do cíngulo;
LM1 (=M1-DMD) – diâmetro mésio-distal do primeiro molar inferior, entre os pontos mais oral e mais aboral do cíngulo. De notar que o estado incompleto deste dente no exemplar de Palmela impossibilita a tomada deste medida. No entanto, foi medido o diâmetro máximo do respectivo alvéolo. Por este motivo, o valor surge no gráfico em linha descontínua. O Diâmetro vestíbulo-lingual do alvéolo não foi considerado.
SDI – comprimento da série dentária inferior, desde o bordo alveolar anterior do primeiro incisivo ao bordo posterior do cíngulo do M1. A ausência da porção posterior do cíngulo impede a correcta tomada da medida. De qualquer modo, optou-se por registar a distância entre o primeiro ponto referido e o bordo alveolar posterior do M1. Tal como na situação anterior, tratando-se de um valor aproximado, fica representado no gráfico em linha descontínua.
C1-M1 – distância entre o bordo alveolar anterior do C1 e o bordo posterior do cíngulo do M1. Também neste caso só foi possível obter uma medição aproximada, tendo como referência o bordo alveolar posterior do M1. Representação no gráfico em linha descontínua.
LM – Comprimento mandibular, desde o ponto mais aboral do processo condilar ao infradental. Conforme o anteriormente referido, as extremidades do processo condilar apresentam-se fracturadas, pelo que a medida registada é ligeiramente inferior à que seria obtida caso a peça estivesse completa. Assim, o valor é igualmente representado no gráfico em linha descontínua.

A leitura do gráfico exposto permite verificar que, como seria de esperar, a presente peça mandibular se enquadra nas dimensões referentes a lince-ibérico. Os valores apresentam-se muito próximos dos máximos referentes a esta espécie e abaixo dos mínimos obtidos em exemplares de lince-boreal, exceptuando o diâmetro vestíbulo-lingual do canino que ultrapassa muito ligeiramente o valor máximo em L. pardina, representando um insignificante aumento da faixa de sobreposição deste parâmetro biométrico entre as duas espécies. Todas as medidas se revelaram superiores às médias para machos das populações ibéricas e, com excepção do P3, também acima das dimensões máximas para fêmeas. De notar que os valores máximos do P3 são semelhantes em ambos os sexos, não possuindo portanto carácter discriminante. Em oposição, o comprimento mandibular (LM), o da série dentária inferior (SDI), o C1-M1 e os diâmetros do canino são todos considerados muito significativos para este efeito, de acordo com o teste t de Student, segundo Garcia-Perea (1991:267). Tal permite concluir que a hemimandíbula de Palmela pertenceu a um exemplar de L. pardina macho de compleição bem desenvolvida.
Na peça em apreço não foram assinaladas marcas evidentes de corte. Existe apenas um pequeno golpe no bordo inferior do ramo mandibular horizontal, com menos de 2 mm de extensão, aparentemente realizado por gume afiado.
Fig. 6: Hemimandíbula esquerda de lince-ibérico macho proveniente do Q. S2-3, U.E. 30.
No dealbar do período moderno o lince era certamente uma espécie com ampla distribuição na Península Ibérica. Era caçado para aproveitamento da pele e devido à sua reputação de predador voraz que matava as criações domésticas e destruía os recursos cinegéticos. Esta imagem negativa sobre o lince atingiu contornos que roçam o absurdo, conforme se pode perceber pela seguinte citação de Galvão et al. (1943-1945:135): “O lince é um dos animais mais cruéis da criação... É sanguinário e cruel, muito além das suas necessidades - doentiamente, como certos sádicos. Mata por prazer, porque o sangue e a agonia das vítimas o deleitam... Se tivesse a corpulência e o poder de um tigre ou de um leão, seria o bicho mais terrível do reino animal... depois do homem é claro.” (sic). Tal fama deve-se sobretudo a alguns dos padrões observados no modo de consumir as presas. No caso dos coelhos é frequente rejeitar parte significativa da presa, particularmente a componente não muscular das pernas traseiras, o estômago, os intestinos e ocasionalmente a cabeça e partes da pele. Dos ungulados consome apenas o músculo, normalmente das zonas mais carnudas dos membros (Castells & Mayo, 1993).
Breve apontamento histórico
É fundamental o conhecimento detalhado do enquadramento político, social e económico relativos ao período de tempo em que se formou o depósito de onde provieram os materiais em estudo. A ponte entre o conhecimento histórico e a investigação estritamente arqueozoológica é crucial, particularmente neste tipo de contextos em que é possível confrontar os testemunhos materiais com os documentais.
A presença de um interessante conjunto de vestígios atribuíveis a fauna selvagem poderá estar em conformidade com alguns aspectos registados da evolução política e social no período abrangido. Recorde-se que durante o Portugal medievo, a realeza não reconhecia ao povo o direito à caça, apesar dos protestos populares (Barros, H., 1885-1922). O problema agudizou-se especialmente no reinado de D. Afonso V, levando este monarca a promulgar um conjunto de leis, posteriormente coligidas no “Livro Vermelho”, respeitantes aos limites das coutadas reais. Esta legislação reconhecia o direito à caça pelas populações e fixava períodos de defeso em relação à captura de algumas espécies. Foram também interditas determinadas técnicas específicas de captura, consideradas particularmente destrutivas (Galvão et al. 1943-1945). De qualquer modo, a maior abertura à possibilidade de qualquer um caçar de modo não furtivo, influenciou seguramente a intensidade da exploração de animais selvagens. Face a estes dados, a quantificação das proporções dos restos de animais selvagens e domésticos e a respectiva comparação com estudos arqueozoológicos de contextos um pouco mais recuados, poderá trazer alguma luz sobre as consequências da legislação adoptada e a sua influência nos hábitos quotidianos em Palmela.
Perspectivas de trabalho
· As tarefas estão de momento interrompidas por falta de disponibilidade do autor. Falta concluir a lavagem dos materiais para prosseguir com as tarefas de identificação taxonómica.
· As contagens terão lugar após a revisão sistemática, de modo a que seja possível averiguar as proporções das espécies representadas. Em cada espécie, vão ser igualmente definidos os índices de representatividade de cada segmento anatómico. Pretende-se realizar as seguintes quantificações:
- NTR = número total de restos;
- NRDT = Número de restos determinados;
- Número de restos de cada espécie;
- NMI = número mínimo de indivíduos de cada espécie.
· Outro aspecto que irá ser considerado consiste na averiguação das idades de abate em algumas das espécies.
· As alterações provocadas nos ossos tais como: marcas de corte, polimentos, afeiçoamentos, incisões e outras serão registadas e analisadas.
· Algumas patologias ósseas foram já detectadas e outras mais poderão surgir ainda. A interpretação das causas que lhes deram origem é mais um dos objectivos que se pretende atingir.
· A medição normalizada das peças permitirá estabelecer comparações biométricas com dados equivalentes publicados, referentes a outros períodos cronológicos. Este tipo de abordagem poderá trazer informações sobre eventuais melhoramentos ou modificações das espécies domésticas.
Bibliografia citada
Antunes, M. T. (1989) – Castor fiber na gruta do Caldeirão. Existência, distribuição e extinção do castor em Portugal. Ciências da Terra, 10:23-40. Universidade Nova de Lisboa.
Amorosi, T. (1989) – A postcranial Guide to Domestic Neo-Natal and Juvenile Mammals. The Identification and Aging of Old World Species. BAR International Series 533.
Baatz, M. & Baatz, M. (1992) – Raças de Cães. Col. Habitat, Ed. Presença.
Barret, P. & Macdonald, D. (1993) – Collins Field Guide – Mammals of Britain and Europe. Harper-Collins Publishers. Versão traduzida “Guias FAPAS – Mamíferos de Portugal e Europa”.
Barros, H. (1885-1922) – História da Administração Pública em Portugal nos séculos XI a XV. Ed. Sá da Costa, 2ª ed., Lisboa. 11 vols.
Boessneck, J. (1969) – Osteological Differences between Sheep (Ovis aries Linné) and Goat (Capra hircus Linné). In Brothwell, D. & Higgs, S. Science in Archaeology. Thames & Hudson. Bristol.
Brewer, D. J. (2001) – The Evolution of the Modern Dog. In Brewer D.; Clark T.; & Phillips, A. Dogs in Antiquity. Aris & Phillips, Warminster.
Cantal, J. (1993) – Estúdio faunistico del yacimiento medieval de Plaza España, Motril (Granada). Arqueologia Medieval, 2:243-260. Ed. Afrontamento.
Cardoso, J. L. (1993a) – Contribuição para o conhecimento dos grandes mamíferos do Plistocénico Superior de Portugal. Oeiras.
Cardoso, J. L. (1993b) – Contribuição para o conhecimento da alimentação em contexto islâmico: estudo dos restos mamalógicos e malacológicos das Mesas do Castelinho (Almodôvar). Arqueologia Medieval, 2:103-124. Ed. Afrontamento.
Cardoso, J. L.; Senna-Martinez, J. & Valera, A. C. (1995/1996) – Aspectos da economia alimentar do Bronze Pleno da Beira Alta: a fauna de grandes mamíferos das “Salas 2 e 20”. Trabalhos de Arqueologia da EAM, 3/4:253-261. Colibri, Lisboa.
Carvalho, A. Rafael (2005) – Intervenção arqueológica no “Mercado Velho” de Palmela – primeiros resultados. al-madan, adenda electrónica, IIª Série (13), em http://almadan.cidadevirtual.pt
Castels, A. & Mayo, M. (1993) – Guia de los Mamíferos en Libertad de España y Portugal. Ed. Pirámide, S. A. – Madrid.
Chaix, L. & Méniel, P. (2001) – Archéozoologie. Les animaux et l’archéologie. Editions Errance, Paris.
Clutton-Brock, J. (1969) – The Origins of the Dog. In Science in Archaeology. A survey of Progress and Research. Ed. Brothwel, D. & Higgs, E. Thames and Hudson. London.
Cohen, A & Sergeantson, D. (1996) – Bird Bones from Archaeological Sites. Archetype Publications Ltd (revised edition), London.
Cornwall. I. (1956) – Bones for the Archaeologist. Phoenix House LTD, London.
Davis, S. (1989) – La Arqueologia de los Animales. Ediciones Bellaterra, S. A., Barcelona.
Desse J., Chaix, L. & Desse-Berset, N. (1986) – “OSTÉO”. Base-réseau de donnés ostéométriques pour l’archéozoology. Centre National de la Recherche Scientifique (CNRS).
Driesch, A. E. von den (1976) – A guide to the Measurement of Animal Bones from Archaeological Sites. Peabody Museum Bulletin, 1. Cambridge.
Galvão, H; Cruz, F. & Montês, A. (1943-1945) – A Caça no Império Português. Ed. “O Primeiro de Janeiro”. Porto.
Garcia-Perea, Rosa (1991) – Variabilidad morfológica del género Lynx Kerr, 1792 (Carnívora: Felidae). Dissertação de doutoramento, Universidad Complutense de Madrid.
Henriques, Francisco da Fonseca (séc. XVIII) – Soccorro Delphico.
Kratochvil, J. & Kratochvil, Z. (1976) – The origin of the domesticated forms of the genus Felis (Mammalia). Zoologické Listy, 25(3): 193-208.
Nobre, A. (1931/40) - Moluscos marinhos de Portugal. Imprensa Portuguesa, Porto.
Izquierdo, E. (1986) – Contribución al Atlas Osteológico de los Teleósteos Ibéricos I. Dentário Y Articular. Ed. Universidad Autónoma de Madrid.
Morey, D. F. (1992) – Size, shape and development in the evolution of the domestic dog. Journal of Archaeological Science, 19:181-204.
Pales & Lambert (1971) – Atlas Ostéologique pour server à l’identification des mammifères du Quaternaire: Herbivores. Ed. du Centre National de la Recherche Scientifique.
Pales & Garcia (1981) – Atlas Ostéologique pour server à l’identification des mammifères du Quaternaire: Carnivores, Homme. Ed. du Centre National de la Recherche Scientifique.
Pires, F.; Cardoso, J. L. & Petrucci-fonseca, F. (2001/2002) – Estudo arqueozoológico dos carnívoros do povoado pré-histórico de Leceia. Estudos Arqueológicos de Oeiras, 10:183-247. Oeiras.
Schmid, E. (1972) – Atlas of Animal Bones. New York: Elsevier Publishing Company.
Macedo, M. & Borges J. (1999) – Conchas Marinhas de Portugal. Verbo.
O’Connor, T (2000) – The archaeology of animal bones. Sutton Publishing.
Reitz E. & Wing, E. (1999) – Zooarchaeology. Cambridge Manuals in Archaeology.
Sabelli, B.; Giannuzi-Savelli, R. & Bedulli, D. (1992) – Catalogo annotato dei molluschi marini del Mediterraneo , vol. 1-3. Società Italiana di Malacologia, Ed. Libreria Naturalistica Bolognese.
Smith, R. N. (1969) – Fusion of ossification centres in the cat. Journal of Small Animal Practice, 10:523-530.
Vasconcelos, R. C. (1998) – Raças de Cães Portugueses. Col. Habitat, Ed. Presença.











0 comentários:
Enviar um comentário